The first Thelocactus to appear in Europe was Thelocactus leucacanthus. This species was discovered by Karwinski near Zimapán, and sent to the München botanical garden around 1830. At that time it was ascribed to the genus Echinocactus, as was the case with most of the globular cacti. The name Thelocactus appeared in the literature many years later, introduced by K. Schumann, who in his 'opus magnum', Gesamtbeschreibung der Kakteen, used it to identify a subgenus of the then mastodontic genus Echinocactus. In it Schumann included a series of species sharing the common character of a stem divided into tubercles, from which the name Thelocactus ('thele' in Greek means nipple).
About thirty years later, Britton and Rose raised Thelocactus to the rank of genus, designating Thelocactus hexaedrophorus as the type species, and including twelve taxa that still belong to it today. After Britton and Rose's work, many other species were added to the original group, both as new species and as combinations. Furthermore, the genus underwent some revisions that modified the taxonomy and therefore the nomenclature of the species involved. Some of the species combined in Thelocactus (T. gielsdorfianus, T. knuthianus, T. saueri, T. viereckii, T. subterraneus) were afterwards included by Backeberg in a related genus, Gymnocactus, today a synonym of Turbinicarpus.
The Eighties saw the description of some new taxa, both at variety and species rank, and, by Anderson, a revision of the genus. In this revision the author included Hamatocactus setispinus in Thelocactus, basing his proposal on the similarities between many of the characters chosen to compare the species Thelocactus bicolor and Hamatocactus setispinus. This new delimitation of Thelocactus totally upset the limits of the genus: the two essential characters on which Britton and Rose had based the genus, i.e. the presence of tubercles, that gave the genus its name, and fruits dehiscent at the base, disappeared, since Hamatocactus setispinus has no tubercles and has fleshy, indehiscent fruits.
Since Anderson's revision, more species have been described (Thelocactus garciae, Thelocactus multicephalus, Thelocactus flavus) and others have been segregated from the genus. Doweld, in 1998, has proposed the separation of Thelocactus conothelos and its varieties from Thelocactus and the inclusion of these taxa in a new genus, all based only on the differences in the micro-morphology of the seed cuticle, that in these species is smooth instead of finely granular. Later, the work by Mosco and Zanovello allowed them to state that Hamatocactus is not congeneric with Thelocactus, and to revise the taxonomic position of a few taxa. In the year 2000 a consistent number of new names has been added to the ones already known, due to the work of some Czechoslovakian authors who have taken the trouble to describe some mere local forms.
The latest reference book to cacti for many years to come is The New Cactus Lexicon, published in 2006 and edited by D. Hunt, N. Taylor and G. Charles on behalf of the International Cactaceae Systematics Group (ICSG). The taxonomic treatment of Thelocactus is more on the conservative side, but there are some choices I do not agree with. The inclusion of Hamatocactus bicolor (syn. Hamatocactus setispinus) in Thelocactus has no grounds due to the relevant differences in body morphology (presence or absence of tubercles) as well as fruit and seed morphology. Thelocactus conothelos ssp. flavus is treated as a synonym of Thelocactus conothelos ssp. aurantiacus, but the two are considerably different for what concerns stem habit and spination. The fact is that T. conothelos ssp. flavus is distinct from any conothelos subspecies for its clustering stems and spination pattern, while T. conothelos ssp. aurantiacus is indistinguishable from the type species if not in flower. I really do not understand the rationale underlaying the synonymization of ssp. flavus and on the contrary the retention of, say, ssp. garciae, which is barely distinguishable from ssp. conothelos. A strong lumping approach was used with the rinconensis complex, where none subspecies was accepted, while for the bicolor complex a different approach was followed and all subspecies were considered "good". This is unintelligible, as to me a T. rinconensis ssp. nidulans is far more distinct from a T. rinconensis than a T. bicolor ssp. bolaensis from a T. bicolor is. The lumping approach did not affect T. hexaedrophorus ssp. lloydii, which is described as having 1-3 central spines and on this basis distinguished from ssp. hexaedrophorus. Interestingly, the original description does not mention the presence of central spines, so where they have found them is a mystery. To conclude the overall taxonomic treatment is incongruent, with some species that were treated in a more conservative way than others were. There is still an error to be pointed out. Thelocactus seeds have a flat or convex relief, but never concave or par-concave as stated in the genus description.
Some nomenclatural changes, proposed by G. F. Matuszewski and S. Hinz, can be found in their monograph Thelocactus, published in 2011. The book is well illustrated with a lot of beautiful pictures depicting any form of the known species. Alas, it is written in Polish and German, which prevents the understanding to the majority of people. The authors revised the nomenclature of the T. rinconensis complex according to their opinion that T. lophothele is identifiable with the plants growing on the northern side of the highway from Saltillo to Monterrey. That both are varieties of the same species is an old idea, but because tha place of origin of lophothele was stated being Chihuahua, where till now no plant matching the description has been found, the name was set apart in the most recent works. If lophothele is considered a good species, say close to rinconensis, nomenclature has to be adjusted, because lophothele has priority over rinconensis, what the authors did. To support their taxonomic point of view, the authors relied on a phenetic analysis based on 50 characters, and here a criticism can be made because they did not said what characters were chosen, and the software used to run the phenetic analysis.
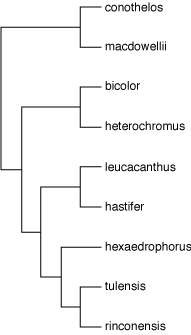
The genomic era has made available new tools for the study of phylogenetic relationships among organisms. Molecular data are now commonly used to study plant lineages at shallow or deep levels, and Cactaceae are no exception. Several studies have been made to test the monophyly of the subfamilies or tribes as well as to resolve intergeneric relationships and to assess the monophyly of extant genera using chloroplast DNA sequence data. Thelocactus belongs to the tribe Cacteae, which is monophyletic, but the limits of its genera are still matters of debate. In their study, Vásquez-Sánchez and coauthors, have found that Ferocactus is polyphyletic because it is grouped in the same clade together with Glandulicactus, Leuchtembergia, Stenocactus and Thelocactus. The authors have suggested considering all these genera as Ferocactus until new data will support them. Nevertheless, this analysis is incomplete, at least for what concerns Thelocactus, because three species, T. buekii, T. lausseri, and T. multicephalus, were not included in the analysis which means that we have to wait for further studies to have a complete understanding of the interspecific relationships in this genus.
History